
Curiosity Without the Commute: Rethinking Research in Rural Aotearoa
Plankton assemblages and spatiotemporal variation | Meadow Simpson
Introduction
Plankton are a diverse group of organisms, including plants (phytoplankton) and animals (zooplankton), that passively drift in the water column [1]. Despite their small size, with the majority of plankton less than 1 mm in size and thus invisible to the unaided human eye, they hold immense value through their role in primary production, carbon sequestration, food provision, and water quality biomarkers [2–7]. That said, the rapid proliferation of certain phytoplankton, such as Skeletonema costatum, termed harmful algal blooms (HABs), has adverse effects on the environment due to oxygen depletion, reduced water quality aesthetics, and toxicity [8]. Monitoring harmful algae is thus essential for environmental, social, and economic factors.
Despite the significance of plankton as detailed above, understanding and predicting the spatial and temporal distribution of plankton species remains difficult due to the various environmental factors at play. These difficulties are pronounced in estuaries in comparison to the open ocean, as they represent a transitional zone between freshwater and marine biomes. These are characterised by fluctuating salinities and a gradual shift in environmental variables [9]. Furthermore, estuarine conditions such as salinity, turbidity, and temperature are shifting with climate change [10–16]. These changes directly and indirectly affect plankton as they influence metabolism, growth, reproduction, morphology, and distribution [17–22].
The current state of knowledge surrounding plankton is dominated by research conducted in the open ocean and thus focuses on marine plankton [23]. Estuaries provide difficult research conditions due to their dynamic nature and project-specific constraints, these primarily being funding and accessibility. Of the research conducted within estuarine environments, it is apparent that the distribution of plankton, both temporally and spatially, is highly variable and context-specific in regards to the scale of variation, whether that be seasonal, annual, or none at all. This research is based in the Hoteo estuary within the Kaipara Harbour, which holds ecological importance as a snapper feeding ground, habitat for shark populations [24], and nesting site for fairy terns [25]. It aims to fill the knowledge gap outlined above and provide a baseline dataset of plankton community composition. This is used to monitor the effects of climate change and inform future management, with the overall goal of protecting the ecosystem and the invaluable services it provides. The two primary research aims of this study are (1) identify whether spatiotemporal variability exists within the plankton community in the Hoteo estuary and (2) investigate the effect of different environmental variables on the plankton community composition and distribution.
Methods
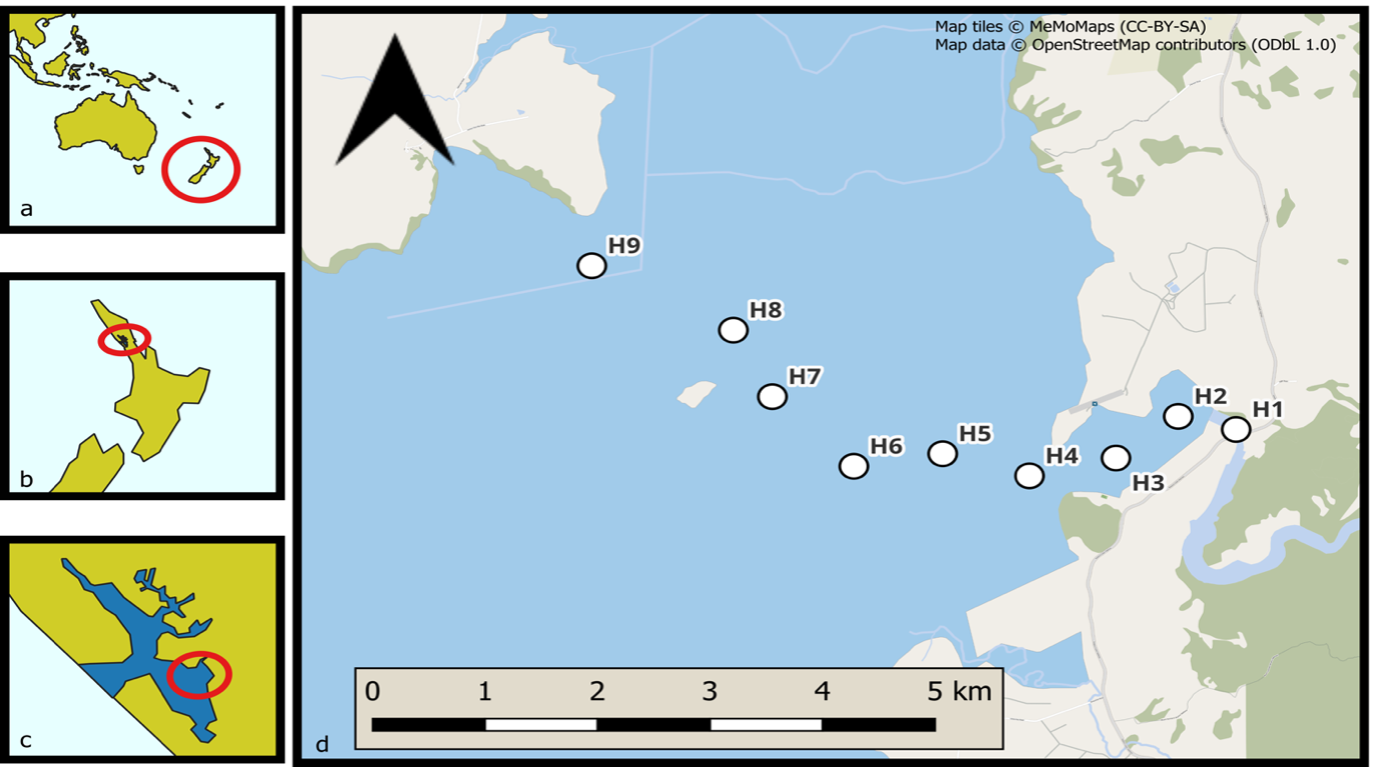
Figure 1: Map of sampling sites along the Hoteo estuary (H1-H9). Samples were taken on February 20th 2024, July 23rd 2024, and February 3rd 2025.
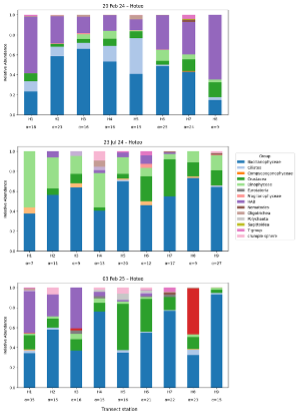
Figure 2: Relative abundance of plankton groups across nine transect sites (H1-H9) along the Hoteo estuary during three sampling periods (20 Feb 2024, 23 Jul 2024, and 03 Feb 2025). Stacked bars represent plankton community composition, with colours corresponding to taxonomic group, and alpha (α) values indicate taxonomic richness.
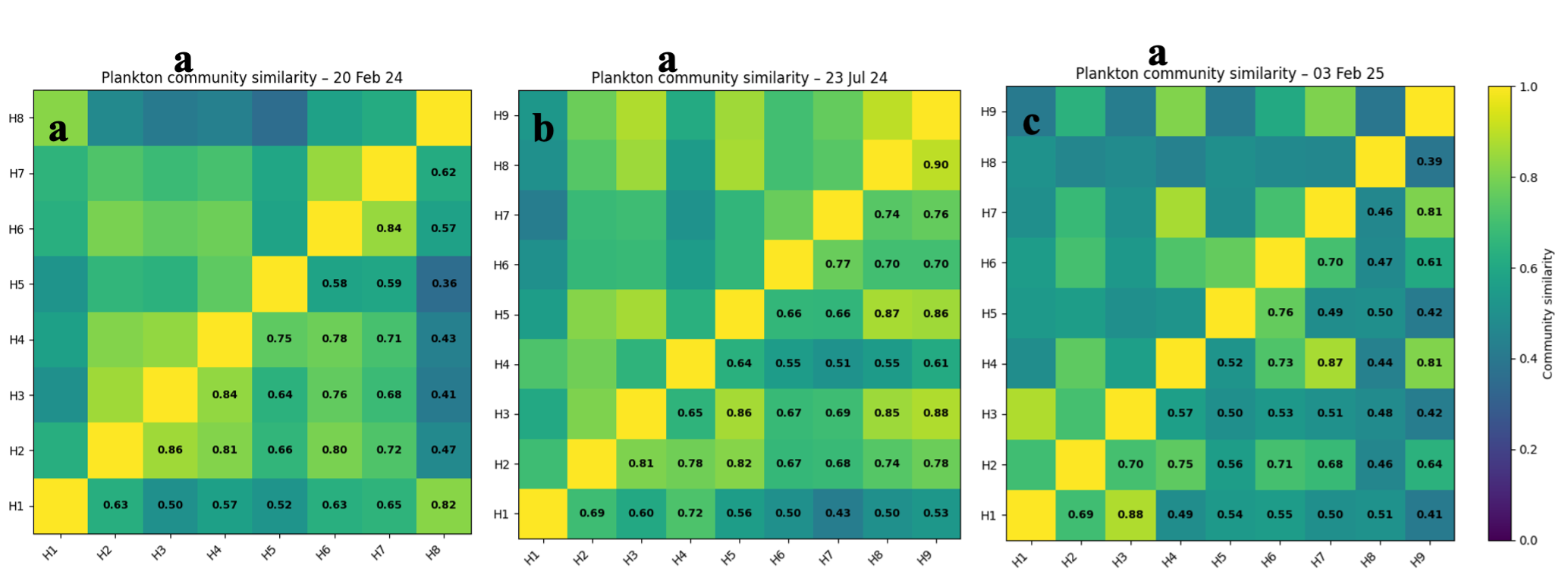
Figure 3: Te Puna o Te Mātauranga marae, Whangārei. A contemporary Māori meeting complex at NorthTec Raumanga campus, used for cultural events, learning workshops, and community gatherings [4].
The research discussed in this article was conducted under the supervision of Anne-Fleur van Leeuwen and utilises the data collected as part of her PhD project, which is currently in preparation for publication (expected 2026). Nine sites, following the natural estuary channel, were sampled along the Hoteo estuary (Figure 1) on February 20th 2024, July 23rd 2024, and February 3rd 2025, representing two austral summer samples (February) and one austral winter sample (July). Each transect site (H1-H9) was sampled on a small boat, primarily, but not always, at high tide. Conductivity, temperature, depth (CTD) and turbidity profiles were collected using an RBR-concerto C.T.D.+ & Seapoint® Turbidity sensor [26]. Chlorophyll a (Chl-a; µg/l) was measured by a C-FLUOR Submersible Probe attached to the RBR-concerto. Samples for surface nutrient levels of ammonium nitrogen (NH₄–N), inorganic nitrogen (NOₓ), and phosphate (PO₄) were collected using a bucket. All equipment was pre rinsed three times before use and samples were filtered on site using a 0.43 µm plastic syringe, chilled and frozen within 12 hours, before analysis in a FIA machine. Plankton samples were collected at the surface using a 16-μm net towed near the surface of the vessel for three minutes. Samples were filtered to contain only individuals between 20-180 μm, stored in 200 ml dark glass bottles and preserved in three drops of neutral Lugol’s iodine.
Imaging was performed using a PlanktoScope [27]. A collection of vignettes were produced and uploaded to EcoTaxa for identification [28]. A total of 56 taxa (Table 2-2) were identified, from 5589 eligible vignettes, based on the World Register of Marine Species (WoRMS) classification index [29], and then grouped into nine classes for visualisation.
Relative abundances, as opposed to absolute taxa abundances, is the primary metric of this report due to the inability to correctly measure the volume sampled for each net tow. The relative abundances of different taxa groups were visualised with bar plots and coloured by class. The taxa compositions were analysed using α-diversity, defined as the number of taxa per sample for each fraction [30]. Salinity, turbidity, tide, and Chl-a were plotted over the initial relative abundance plots to visualise any relationships between these environmental factors and variation in plankton assemblages. A Bray-Curtis dissimilarity index was performed to measure how the relative abundance differed between sites (H1-H9) and sampling periods (20.02.2024, 23.07.2024, 03.02.2025). The results were converted to a similarity measure (1 – Bray-Curtis dissimilarity) and visualised with heat maps. The effect of environmental conditions on plankton community composition was analysed using a redundancy analysis (RDA). It was performed using the rda () function [31] from the scikit-bio Python package [32]. Pearson correlation coefficients [33] were then calculated to obtain correlation significance, defined by having a p-value less than 0.05.
Results
Plankton community composition within the Hoteo estuary exhibited pronounced spatial and temporal variability across the three sampling periods (Figure 2). In February 2024, Bacillariophyceae displayed a clear bell-shaped distribution along the estuarine gradient, increasing from 23% at the upstream site (H1) to a peak of 66% at H3 before declining to 15% at the most marine site (H8). In contrast, harmful algal bloom (HAB) plankton, Skeletonema costatum, showed an inverse pattern, contributing over half of the assemblage at both ends of the estuary (56% at H1, 65% at H8) and displaying minimum values (11–17%) at mid-estuary sites (H3–H5). Ciliates peaked at H5 (36%), while Crustacea, Dinophyceae, Oligotrichea, and Nematodes were comparatively minor components. In July 2024, community composition was dominated by Bacillariophyceae, with the relative abundance fluctuating between 38% (H1) and 73% (H8). Dinophyceae were prominent in the upper estuary (H1–H5), contributing 17–56% of total abundance and dominating at the lowest salinity (7.5 PSU). Crustacea were most abundant downstream, particularly at H6 (25%) and H7 (37%). HAB plankton were negligible during this winter period. February 2025 showed substantial spatial heterogeneity whereby HAB plankton dominated at H1 (42%) and H3 (41%), whereas Bacillariophyceae dominated multiple sites (H2, H4, H7, H9), and reached up to 93% at H9. Crustacea comprised approximately half of the assemblage at H5 (46%) and, together with Bacillariophyceae, dominated H6. Foraminifera appeared for the first time and dominated H8 (46%).
Alpha diversity ranged from 7 to 35 taxa across all periods, with no consistent spatial trend. Both the lowest (7 in July 2024) and highest (35 in February 2025) values occurred at H1. Mean diversity was lowest in July 2024 (13.9) compared to approximately 20 during both summer sampling periods.
Projects such as Tiakina ngā Tini Taonga o Tatou Awaawa, coordinated by Ngā Kaitiaki o Ngā Kai Māori (KNONKM), which samples 16 hapori wai sites to extend understanding of waterways significant to hapū and support decisions on restoration actions, from fencing and planting to fish surveys [5]. Supported by funding streams like the Te Mana o te Wai Fund, these initiatives generate data that feeds directly into regional planning and policy conversations [6].
These projects are long‑running, relational, and centred around place, designed by and for the communities who live alongside the ecosystems they care about. They show that research doesn’t require a city, it requires time, trust, and taking local knowledge seriously.
Science in Your Socks
Doing research in a rural setting doesn’t mean waiting for a lab door to swing open. It often means starting where you are, with whatever’s around you. Think of it as “science in your socks”: paying attention to the world while you’re still standing in it.
Environmental noticing
Keep an eye on the early lambs, the flowers that pop up before you’ve finished your coffee, or how the water in the stream changes after a heavy rain. These observations are probably already happening without even thinking about it. Jotting them down is the first step toward turning everyday curiosity into data.
Table 1: Significant correlations (p<0.05) between plankton groups along the Hoteo estuary and environmental variables (salinity, temperature, and chlorophyll-a). Positive values indicate a positive significant correlation and negative values indicate a negative significant correlation. Values in red denote the strongest relationships in instances where there are significant correlations with numerous environmental variables.
Salinity influenced several taxa (Table 1). HAB plankton occurred only during summer at salinities of 25–35 PSU, while Foraminifera were restricted to the highest salinity (>35 PSU). Crustacea were positively correlated with salinity and increased along the winter salinity gradient, particularly at H6. Dinophyceae were negatively correlated with temperature and salinity and dominated at low salinity. Bacillariophyceae occurred across the full salinity range (23–80%). Turbidity patterns varied seasonally. In July 2024, high turbidity (>30 NTU) characterised upper sites and coincided with Dinophyceae dominance at H1 (55.4 NTU). In February 2025, turbidity peaked at H2–H3 (~70 NTU) but was negligible elsewhere; HAB dominance at H8 coincided with elevated turbidity (~30 NTU). Overall, Bacillariophyceae were abundant in both turbid and clear waters, while Crustacea were most abundant under low turbidity. Chlorophyll-a concentrations were generally low (<4 µg L⁻¹) with isolated spikes, but no significant correlations were detected between chlorophyll-a and plankton groups. Significant correlations (p<0.05) were identified between temperature and ciliates/HAB plankton (positive), and between temperature/salinity and Coscinodiscaceae/Dinophyceae (negative). Overall, seasonal shifts, salinity gradients, and site-specific heterogeneity structured plankton assemblages within the estuary.
Discussion
This study examined spatial and temporal variation in plankton community composition within the Hoteo Estuary and evaluated the environmental factors influencing these patterns. Overall, the results support the hypothesis that plankton assemblages vary both spatially along the estuarine gradient and temporally across sampling periods. Several plankton groups displayed clear differences in relative abundance between transect stations and seasons, reflecting the complex interactions between environmental variables such as salinity, temperature, and turbidity.
Bacillariophyceae were present across all transect stations and sampling periods, although their relative abundance varied. This widespread occurrence likely reflects the taxonomic diversity within this group, which includes numerous diatom genera capable of occupying a wide range of habitats [23]. The dominance of Bacillariophyceae is consistent with previous research showing that diatoms commonly dominate estuarine plankton communities [34, 35]. During the February 2024 sampling period, an inverse relationship was observed between Bacillariophyceae and the harmful algal bloom (HAB) species Skeletonema costatum. At sites where Skeletonema costatum was dominant, Bacillariophyceae abundance was generally lower, and vice versa. Harmful algal species such as Skeletonema costatum often gain a competitive advantage under favourable environmental conditions enabling them to rapidly proliferate and form blooms [36].
Ciliates were also widely distributed across the estuary during February 2024, with particularly high relative abundance at station H5, although they were absent at H7. Interestingly, the Bray–Curtis similarity index indicated that plankton communities at H1 and H8 were relatively similar despite their positions at opposite ends of the estuary, suggesting that factors other than spatial distance may influence plankton community structure.
Clear seasonal differences were observed between summer and winter sampling periods. In July 2024, during the austral winter, Ciliates and Skeletonema costatum were replaced by Dinophyceae in the estuarine transect stations (H1–H5), and by crustaceans in the marine stations (H6–H9). This shift partially aligns with previously documented seasonal successions in temperate marine environments, where ciliates tend to dominate during late winter and early spring, while dinoflagellates become more prevalent in late spring and summer [37]. However, the seasonal pattern observed in this study occurred in the opposite order, with ciliates dominating during summer and dinoflagellates during winter. This apparent contradiction may be explained by the environmental conditions present in the Hoteo Estuary during the winter sampling period. High freshwater input created strong stratification, which may have favoured the growth of dinoflagellates during winter.
In addition to seasonal differences, interannual variability was observed between the two summer sampling periods. In February 2025, Skeletonema costatum was confined to the upper estuarine stations (H1–H3), while crustaceans were abundant at stations H5 and H6, and foraminifera dominated at the marine station H8. Previous studies have reported higher zooplankton densities near estuary mouths, which is consistent with the high crustacean abundance observed at these stations [38]. A major rainfall event occurred prior to the February 2025 sampling, which may have influenced plankton distribution and contributed to the observed differences in community composition. However, this pattern contrasts with previous research suggesting that Skeletonema species often dominate during flood events [39].
Alpha diversity also varied across space and time with the highest and lowest diversity occurring at station H1 in February 2025 and July 2024, respectively. Species richness is expected to increase with salinity, as more species are capable of tolerating marine conditions than freshwater environments [40]. While this pattern explains the low diversity recorded at H1 during the winter sampling period, it does not explain the high diversity recorded at the same location during the February 2025 sampling. This discrepancy suggests that additional environmental factors influence species richness in the estuary. Furthermore, models linking estuarine diversity to salinity gradients have largely been developed for benthic or mobile organisms [9]. These may not be fully applicable to planktonic species, which are transported with tidal currents and therefore may not experience prolonged exposure to specific salinity conditions [41].
Several environmental variables were significantly correlated with the relative abundance of particular plankton groups. Temperature, salinity, and turbidity each influenced different taxa. Ciliates and Skeletonema costatum displayed positive correlations with temperature, while Coscinodiscaceae and Dinophyceae showed negative correlations. Higher temperatures can increase metabolic and growth rates in phytoplankton, helping explain the greater abundance of Skeletonema costatum during summer sampling periods [42]. However, this species is also known to tolerate a wide temperature range, suggesting that other factors also contribute to bloom formation, including nutrient availability, ocean circulation patterns, and anthropogenic disturbances [43–46].
Salinity was positively correlated with Crustacean abundance, consistent with previous findings that many Crustacean zooplankton prefer higher salinity conditions [38]. Conversely, Dinophyceae showed a negative correlation with salinity despite their documented tolerance for salinity variation [37], indicating that additional environmental factors influence their distribution. Ciliates, which are more sensitive to salinity changes, were absent during the July 2024 sampling period when a strong salinity gradient was present. Foraminifera were most abundant in high salinity conditions, consistent with previous studies showing that their growth and reproduction decline as salinity decreases due to the dissolution of their calcareous shells [47, 48].
Turbidity also influenced plankton distribution, though its effects varied among taxa. Ciliates were negatively correlated with turbidity, while Dinophyceae showed a positive correlation. Peaks in crustacean abundance coincided with lower turbidity levels, whereas harmful algal blooms occurred during more turbid conditions. Increased turbidity can directly reduce plankton abundance by inhibiting feeding and reproduction [49–51], but it can also have indirect effects by limiting light availability, reducing primary production, and altering predator–prey interactions [50, 52, 53]. Some dinoflagellate species may possess adaptations that allow them to outcompete other plankton in turbid conditions [54, 55].
The findings of this study also have implications for understanding how climate change may affect estuarine plankton communities. Climate projections suggest that many estuarine systems are experiencing warming, acidification, and freshening [56, 57]. If similar trends occur within the Kaipara Harbour, species positively correlated with salinity, such as crustaceans, may decline, while species negatively correlated with salinity, such as Dinophyceae, may increase. Rising temperatures may also favour harmful algal blooms and alter their seasonal timing and duration [58, 59]. Additionally, climate-driven changes in rainfall may increase turbidity and nutrient runoff, further influencing plankton dynamics [60, 61]. These environmental changes may trigger cascading ecological effects due to competition and trophic interactions within plankton communities.
Despite these insights, the study has several limitations. The use of the Planktoscope and EcoTaxa provided an efficient and affordable method for plankton analysis but limited observations to organisms within the 20–400 µm size range. Automated identification methods can also introduce taxonomic errors, particularly when distinguishing morphologically similar species [62]. To simplify analysis, plankton were grouped into broad taxonomic classes, which may have obscured species-specific environmental responses. Additionally, sampling occurred on single days rather than continuously, meaning that the results represent short-term snapshots rather than long-term trends. Sampling was also conducted primarily at high tide, which may not capture variations that occur across tidal cycles.
Future research should therefore focus on long-term monitoring programs that collect continuous time-series data. Such data would improve the ability to detect seasonal and interannual patterns in plankton communities. Greater taxonomic expertise during identification could also reduce classification errors. Finally, further investigation of ecological interactions such as predation and competition would help clarify how environmental changes influence plankton community structure.
Conclusion
Plankton communities within the Hoteo Estuary exhibit clear spatiotemporal variation influenced by multiple environmental variables. While salinity, temperature, and turbidity explain some of the observed patterns, estuarine plankton dynamics remain highly context-specific and complex. Predicting how climate change will affect these communities remains challenging, highlighting the importance of continued monitoring and research to support effective estuarine management and protect the ecosystem services these environments provide.
[1] G. Hays, A. Richardson, and C. Robinson, “Climate change and marine plankton,” Trends Ecol. Evol., vol. 20, no. 6, pp. 337–344, Jun. 2005, doi: 10.1016/j.tree.2005.03.004.
[2] M. Grigoratou et al., “The immeasurable value of plankton to humanity,” BioScience, vol. 75, no. 9, pp. 706–721, Sep. 2025, doi: 10.1093/biosci/biaf049.
[3] K. Brander, Plankton and Fisheries, vol. 1. Oxford University Press, 2017. doi: 10.1093/oso/9780199233267.003.0008.
[4] P. A. Díaz and G. Álvarez, “Effects of Microalgal Blooms on Aquaculture and Fisheries,” Fishes, vol. 8, no. 9, p. 461, Sep. 2023, doi: 10.3390/fishes8090461.
[5] G. Beaugrand, Plankton Biodiversity and Biogeography, vol. 1. Oxford University Press, 2017. doi: 10.1093/oso/9780199233267.003.0002.
[6] A. Leone, R. Lecci, M. Durante, F. Meli, and S. Piraino, “The Bright Side of Gelatinous Blooms: Nutraceutical Value and Antioxidant Properties of Three Mediterranean Jellyfish (Scyphozoa),” Mar. Drugs, vol. 13, no. 8, pp. 4654–4681, Jul. 2015, doi: 10.3390/md13084654.
[7] G. R. Barroso, L. N. L. Gomes, and S. C. Oliveira, “Planktonic communities as indicators of water quality in a tropical reservoir,” Environ. Manage., vol. 75, no. 6, pp. 1571–1588, Jun. 2025, doi: 10.1007/s00267-025-02157-7.
[8] E. Granéli and J. T. Turner, Eds., Ecology of Harmful Algae, vol. 189. in Ecological Studies, vol. 189. Berlin, Heidelberg: Springer Berlin Heidelberg, 2006. doi: 10.1007/978-3-540-32210-8.
[9] K. Muylaert, K. Sabbe, and W. Vyverman, “Changes in phytoplankton diversity and community composition along the salinity gradient of the Schelde estuary (Belgium/The Netherlands),” Estuar. Coast. Shelf Sci., vol. 82, no. 2, pp. 335–340, Apr. 2009, doi: 10.1016/j.ecss.2009.01.024.
[10] A. Arias-Ortiz et al., “A marine heatwave drives massive losses from the world’s largest seagrass carbon stocks,” Nat. Clim. Change, vol. 8, no. 4, pp. 338–344, Apr. 2018, doi: 10.1038/s41558-018-0096-y.
[11] N. Chen, M. D. Krom, Y. Wu, D. Yu, and H. Hong, “Storm induced estuarine turbidity maxima and controls on nutrient fluxes across river-estuary-coast continuum,” Sci. Total Environ., vol. 628–629, pp. 1108–1120, Jul. 2018, doi: 10.1016/j.scitotenv.2018.02.060.
[12] T. L. Delworth and F. Zeng, “The Impact of the North Atlantic Oscillation on Climate through Its Influence on the Atlantic Meridional Overturning Circulation,” J. Clim., vol. 29, no. 3, pp. 941–962, Feb. 2016, doi: 10.1175/JCLI-D-15-0396.1.
[13] I. García-Moreiras, V. Pospelova, S. García-Gil, and C. Muñoz Sobrino, “Climatic and anthropogenic impacts on the Ría de Vigo (NW Iberia) over the last two centuries: A high-resolution dinoflagellate cyst sedimentary record,” Palaeogeogr. Palaeoclimatol. Palaeoecol., vol. 504, pp. 201–218, Sep. 2018, doi: 10.1016/j.palaeo.2018.05.032.
[14] J. R. Tweedley, C. S. Hallett, R. M. Warwick, K. R. Clarke, and I. C. Potter, “The hypoxia that developed in a microtidal estuary following an extreme storm produced dramatic changes in the benthos,” Mar. Freshw. Res., vol. 67, no. 3, pp. 327–341, Jun. 2015, doi: 10.1071/MF14216.
[15] G. Wang and W. Cai, “Climate-change impact on the 20th-century relationship between the Southern Annular Mode and global mean temperature,” Sci. Rep., vol. 3, no. 1, p. 2039, Jun. 2013, doi: 10.1038/srep02039.
[16] Intergovernmental Panel On Climate Change, “Changing Ocean, Marine Ecosystems, and Dependent Communities,” in The Ocean and Cryosphere in a Changing Climate: Special Report of the Intergovernmental Panel on Climate Change, 1st ed., Cambridge University Press, 2022, pp. 447–588. doi: 10.1017/9781009157964.
[17] D. Albini et al., “Warming alters plankton body-size distributions in a large field experiment,” Commun. Biol., vol. 8, no. 1, p. 162, Feb. 2025, doi: 10.1038/s42003-024-07380-2.
[18] M. J. Cabrerizo and E. Marañón, “Temperature fluctuations in a warmer environment: impacts on microbial plankton,” Fac. Rev., vol. 10, p. 9, 2021, doi: 10.12703/r/10-9.
[19] C. Fernández-González, G. A. Tarran, N. Schuback, E. M. S. Woodward, J. Arístegui, and E. Marañón, “Phytoplankton responses to changing temperature and nutrient availability are consistent across the tropical and subtropical Atlantic,” Commun. Biol., vol. 5, no. 1, p. 1035, Sep. 2022, doi: 10.1038/s42003-022-03971-z.
[20] T. Soulié, F. Vidussi, J. Courboulès, S. Mas, and B. Mostajir, “Metabolic responses of plankton to warming during different productive seasons in coastal Mediterranean waters revealed by in situ mesocosm experiments,” Sci. Rep., vol. 12, no. 1, p. 9001, May 2022, doi: 10.1038/s41598-022-12744-x.
[21] A. Toseland et al., “The impact of temperature on marine phytoplankton resource allocation and metabolism,” Nat. Clim. Change, vol. 3, no. 11, pp. 979–984, Nov. 2013, doi: 10.1038/nclimate1989.
[22] X. Ding et al., “Phytoplankton Communities Miniaturization Driven by Extreme Weather in Subtropical Estuary under Climate Changes,” Water Res., vol. 245, p. 120588, Oct. 2023, doi: 10.1016/j.watres.2023.120588.
[23] R. Adhiambo, P. K. Mensah, and E. Acheampong, “Widespread Geographical Disparities in Phytoplankton Ecology Research in the Face of Climate Change: A Review,” Water, vol. 15, no. 24, p. 4288, Dec. 2023, doi: 10.3390/w15244288.
[24] M. Francis, W. Lyon, E. Jones, P. Notman, D. Parkinson, and C. Getzlaff, Rig nursery grounds in New Zealand: a review and survey. Wellington [N.Z.]: Ministry for Primary Industries, 2012.
[25] K. Baird, S. Ismar, D. Wilson, S. Plowman, R. Zimmerman, and M. Bellingham, “Sightings of New Zealand fairy tern (Sternula nereis davisae) in the Kaipara Harbour following nest failure,” Notornis, vol. 60, pp. 183–185, Jun. 2013, doi: 10.63172//243958iaeomz.
[26] RBR Global, “RBRDuo3 & RBRConcerto3 | C.T, C.T.D, C.T.D++ | Ocean CTD,” RBR, Mar. 20, 2026. https://rbr-global.com/products/standard-loggers/rbrduo-ct/
[27] T. Pollina et al., “PlanktoScope: Affordable Modular Quantitative Imaging Platform for Citizen Oceanography,” Front. Mar. Sci., vol. 9, p. 949428, Jul. 2022, doi: 10.3389/fmars.2022.949428.
[28] Picheral M, Colin S, and Irisson J-O, EcoTaxa, a tool for the taxonomic classification of images. [Online]. Available: http://ecotaxa.obs-vlfr.fr
[29] “WORMS - World Register of Marine Species.” https://www.marinespecies.org/index.php
[30] R. H. Whittaker, “Vegetation of the Siskiyou Mountains, Oregon and California,” Ecol. Monogr., vol. 30, no. 3, pp. 279–338, 1960, doi: 10.2307/1943563.
[31] Pierre Legendre and Louis Legendre, Numerical Ecology, 3rd ed., vol. 24. 2012. Accessed: Feb. 10, 2026. [Online]. Available: https://shop.elsevier.com/books/numerical-ecology/legendre/978-0-444-53868-0
[32] Matthew Aton, Daniel McDonald, and Jorge Cañardo Alastuey, Scikit-bio: a fundamental Python library for biological omic data analysis. (2025).
[33] K. Pearson, “Note on Regression and Inheritance in the Case of Two Parents,” Proc. R. Soc. Lond., vol. 58, pp. 240–242, 1895.
[34] J. Hasan et al., “Phytoplankton Community in Relation to Environmental Variables in the Tidal Mangrove Creeks of the Pasur River Estuary, Bangladesh,” Conservation, vol. 2, no. 4, pp. 587–612, Oct. 2022, doi: 10.3390/conservation2040039.
[35] H. Yoo, C. W. Ji, and I.-S. Kwak, “Long-term analysis of Bacillariophyta assemblages in Korean estuaries: effects of salinity gradients and environmental factors (2008–2019),” Hydrobiologia, vol. 853, no. 2, pp. 395–411, Jan. 2026, doi: 10.1007/s10750-025-05937-8.
[36] S. Chakraborty and U. Feudel, “Harmful algal blooms: combining excitability and competition,” Theor. Ecol., vol. 7, no. 3, pp. 221–237, Aug. 2014, doi: 10.1007/s12080-014-0212-1.
[37] A. Calbet, “On the ecological duality between ciliates and dinoflagellates across marine ecosystems,” J. Plankton Res., vol. 47, no. 5, p. fbaf049, Sep. 2025, doi: 10.1093/plankt/fbaf049.
[38] R. Adhiambo, P. K. Mensah, E. Acheampong, and E. A. Krampah, “Plankton assemblages in a tropical West African estuary,” Aquat. Sci., vol. 86, no. 3, p. 70, May 2024, doi: 10.1007/s00027-024-01085-8.
[39] P. Y. Guo, H. T. Shen, and J. H. Wang, “Species diversity, community structure and distribution of phytoplankton in the Changjiang estuary during dry and flood periods,” J. Mar. Biol. Assoc. U. K., vol. 94, no. 3, pp. 459–472, May 2014, doi: 10.1017/S0025315413001641.
[40] I. D. S. Orizar and A. M. Lewandowska, “Interspecific trait variability and plasticity of the Baltic Sea phytoplankton species along a salinity gradient,” J. Plankton Res., vol. 47, no. 3, p. fbaf015, Apr. 2025, doi: 10.1093/plankt/fbaf015.
[41] M. J. Attrill, “A testable linear model for diversity trends in estuaries,” J. Anim. Ecol., vol. 71, no. 2, pp. 262–269, 2002, doi: 10.1046/j.1365-2656.2002.00593.x.
[42] H. Li, T. Xu, J. Ma, F. Li, and J. Xu, “Physiological responses of Skeletonema costatum to the interactions of seawater acidification and the combination of photoperiod and temperature,” Biogeosciences, vol. 18, no. 4, pp. 1439–1449, Feb. 2021, doi: 10.5194/bg-18-1439-2021.
[43] N. Yu. Shoman and A. I. Akimov, “Features of Temperature Adaptation of Phaeodactylum tricornutum, Nitzschia sp., and Skeletonema costatum (Bacillariophyceae) under Different Light Conditions,” Dokl. Biol. Sci., vol. 506, no. 1, pp. 256–263, Oct. 2022, doi: 10.1134/S0012496622050155.
[44] Y. Suzuki and M. Takahashi, “Growth Responses of Several Diatom Species Isolated from Various Environments to Temperature,” J. Phycol., vol. 31, no. 6, pp. 880–888, 1995, doi: 10.1111/j.0022-3646.1995.00880.x.
[45] D. G. Borkman and T. J. Smayda, “Gulf Stream position and winter NAO as drivers of long-term variations in the bloom phenology of the diatom Skeletonema costatum ‘species-complex’ in Narragansett Bay, RI, USA,” J. Plankton Res., vol. 31, no. 11, pp. 1407–1425, Nov. 2009, doi: 10.1093/plankt/fbp072.
[46] W. Hou et al., “Characteristics of Harmful Algal Species in the Coastal Waters of China from 1990 to 2017,” Toxins, vol. 14, no. 3, Feb. 2022, doi: 10.3390/toxins14030160.
[47] N. Sahoo, S. M. Saalim, A. Matul, R. Mohan, A. Tikhonova, and N. Kozina, “Planktic Foraminiferal Assemblages in Surface Sediments From the Subpolar North Atlantic Ocean,” Front. Mar. Sci., vol. 8, Feb. 2022, doi: 10.3389/fmars.2021.781675.
[48] R. Saraswat, M. Kouthanker, S. R. Kurtarkar, R. Nigam, S. W. A. Naqvi, and V. N. Linshy, “Effect of salinity induced pH/alkalinity changes on benthic foraminifera: A laboratory culture experiment,” Estuar. Coast. Shelf Sci., vol. 153, pp. 96–107, Feb. 2015, doi: 10.1016/j.ecss.2014.12.005.
[49] R. C. Hart, “Zooplankton feeding rates in relation to suspended sediment content: potential influences on community structure in a turbid reservoir,” Freshw. Biol., vol. 19, no. 1, pp. 123–139, Feb. 1988, doi: 10.1111/j.1365-2427.1988.tb00334.x.
[50] J. D. Jack and J. J. Gilbert, “The effect of suspended clay on ciliate population growth rates,” 1993, Accessed: Feb. 04, 2026. [Online]. Available: https://onlinelibrary.wiley.com/doi/10.1111/j.1365-2427.1993.tb00773.x
[51] S. Jones, N. K. Carrasco, and R. Perissinotto, “Turbidity effects on the feeding, respiration and mortality of the copepod Pseudodiaptomus stuhlmanni in the St Lucia Estuary, South Africa,” J. Exp. Mar. Biol. Ecol., vol. 469, pp. 63–68, Aug. 2015, doi: 10.1016/j.jembe.2015.04.015.
[52] P. M. D’Costa, A. Chandrashekar Anil, J. S. Patil, S. Hegde, M. S. D’Silva, and M. Chourasia, “Dinoflagellates in a mesotrophic, tropical environment influenced by monsoon,” Estuar. Coast. Shelf Sci., vol. 77, no. 1, pp. 77–90, Mar. 2008, doi: 10.1016/j.ecss.2007.09.002.
[53] R. Jones et al., “Underwater Light Characteristics of Turbid Coral Reefs of the Inner Central Great Barrier Reef,” Front. Mar. Sci., vol. 8, Nov. 2021, doi: 10.3389/fmars.2021.727206.
[54] C. Fan and P. M. Glibert, “Effects of light on nitrogen and carbon uptake during a Prorocentrum minimum bloom,” Harmful Algae, vol. 4, no. 3, pp. 629–641, Mar. 2005, doi: 10.1016/j.hal.2004.08.012.
[55] F. Hilaluddin, F. M. Yusoff, and T. Toda, “Shifts in Diatom Dominance Associated with Seasonal Changes in an Estuarine-Mangrove Phytoplankton Community,” J. Mar. Sci. Eng., vol. 8, no. 7, Jul. 2020, doi: 10.3390/jmse8070528.
[56] F. Estrada, D. Kim, and P. Perron, “Spatial variations in the warming trend and the transition to more severe weather in midlatitudes,” Sci. Rep., vol. 11, no. 1, p. 145, Jan. 2021, doi: 10.1038/s41598-020-80701-7.
[57] P. Pfleiderer et al., “Considerable yet contrasting regional imprint of circulation change on summer temperature trends across the Northern hemisphere mid-latitudes,” Weather Clim. Dyn., vol. 7, no. 1, pp. 89–108, Jan. 2026, doi: 10.5194/wcd-7-89-2026.
[58] M. Edwards and A. J. Richardson, “Impact of climate change on marine pelagic phenology and trophic mismatch,” Nature, vol. 430, no. 7002, pp. 881–884, Aug. 2004, doi: 10.1038/nature02808.
[59] L. Gao, Y. Xiong, F.-X. Fu, D. A. Hutchins, K. Gao, and G. Gao, “Marine heatwaves alter competition between the cultured macroalga Gracilariopsis lemaneiformis and the harmful bloom alga Skeletonema costatum,” Sci. Total Environ., vol. 947, p. 174345, Oct. 2024, doi: 10.1016/j.scitotenv.2024.174345.
[60] C.-S. Lee, Y.-C. Lee, and H.-M. Chiang, “Abrupt state change of river water quality (turbidity): Effect of extreme rainfalls and typhoons,” Sci. Total Environ., vol. 557–558, pp. 91–101, Jul. 2016, doi: 10.1016/j.scitotenv.2016.02.213.
[61] M. Postacchini, A. J. Manning, J. Calantoni, J. P. Smith, and M. Brocchini, “A storm driven turbidity maximum in a microtidal estuary,” Estuar. Coast. Shelf Sci., vol. 288, p. 108350, Aug. 2023, doi: 10.1016/j.ecss.2023.108350.
[62] Y. G. Lee, “Mapping Plankton Variability Across the Tasman Front: Insights from Planktoscope Imaging.”

Meadow is a sixth year student with a particular interest in sustainability and climate change adaptation. Last year she completed her Honours in Marine Science which focused on testing the dissipative capacity of a temporary structure to mitigate coastal erosion.