
Research Heroes and Data Woes: The Value and Limitations of Research Databases; A Case Study Using Traits of Recently Naturalised Plant Species in Aotearoa
Databases and Biosecurity| Amie Cummack
Introduction
The value of databases for research is widely acknowledged. Open access data can be interpreted and analysed by others if accessible [1], and can be used to identify and investigate patterns, develop and test hypotheses, and build knowledge [2], [3]. Databases allow sharing of the time and effort invested in generating data and a larger return on the resources invested, with greater sharing potential for international databases [1], [4]. However, issues exist with data collection and sharing. Data entered are frequently collected without an explicit intent to share, vary in quality, format, and units, and may not be standardised. Furthermore, combining data from multiple sources can exacerbate issues associated with accuracy [5]–[7]. Data openness can be limited by infrastructure constraints, a lack of incentives for researchers, and legal and ethical issues [1]. The applications of databases require a sufficient set of open data [7], and incompleteness can impact data representativeness, inference accuracy, and the usefulness of applications [2], [8].
Databases can be highly useful for conservation and pest management, for example, improving understanding and management of emerging weeds in Aotearoa [9]. Environmental weeds pose a significant threat to native plant diversity [10]. To be effective and time- and resource-efficient, weed management must occur at the early stages of invasion; however, a large source pool of introduced species means empirical research on each species’ potential impacts is not available [11], [12]. Therefore, database use is valuable here to assess traits and the potential risk of introduced plants. This case study aims to assess the completeness of trait data (fruit type, dispersal mode, and shade tolerance) gathered using global and local plant trait databases for the 70 recently naturalised plant species in Aotearoa.
Glossary
Casual-occurring: reproducing only close to cultivated parent plants, or infrequently in the wild
Environmental weeds: weeds of natural ecosystems. They must be fully naturalised, exist in natural ecosystem areas, and have moderate to massive impacts.
Homotypic synonyms: different binomial/scientific names tied to the same type specimen (physical reference sample)
Naturalised: forming self-sustaining wild populations
Open access: available online for use by anyone without cost or legal barriers.
Species concepts: criteria for defining a species, e.g. biological species concept, ecological species concept.
Figure 1: Global Databases [13].
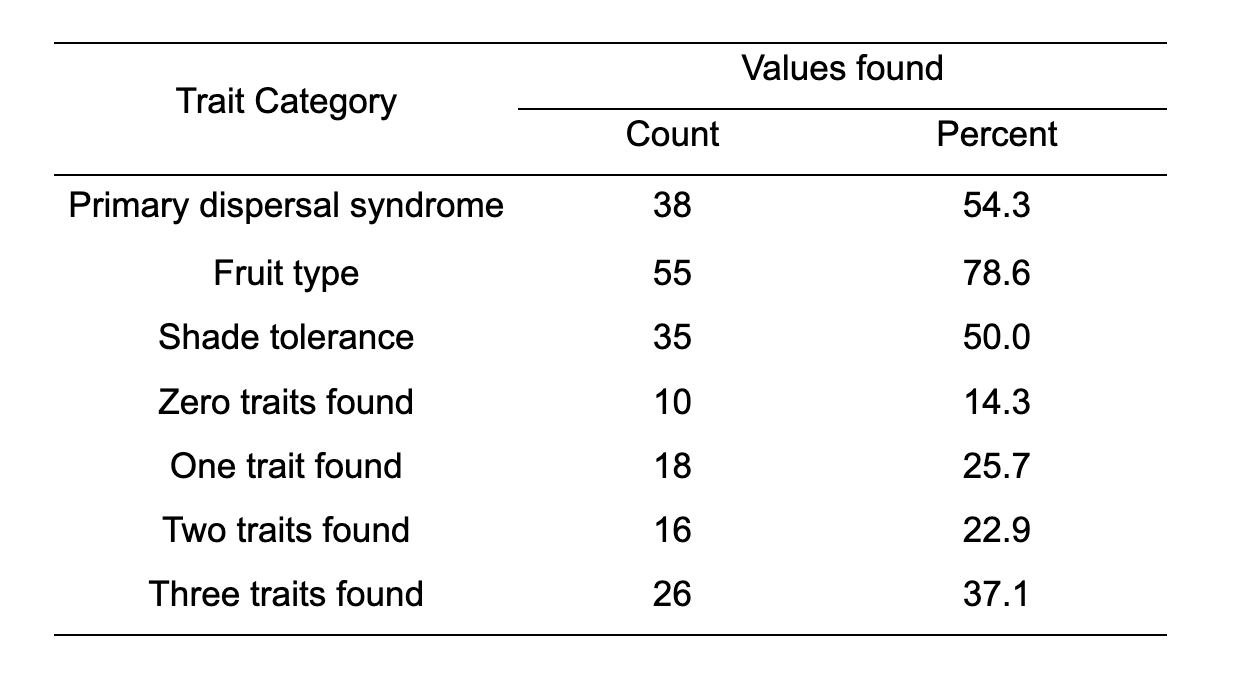
Table 1: Proportion of values explicitly found for traits of the 70 recently naturalised species in Aotearoa using databases, flora, and other online sources. Values inferred using other members of a given genus were excluded.
Discussion
Relatively low data completeness for the 70 target species means these results may not be representative of the traits of recently naturalised plant species in Aotearoa. This appears to be consistent with expectations for database completeness. While data sharing in ecology is increasing, and databases such as TRY and AusTraits have impressive and growing coverage [15], [18], low levels of data are open access. For example, in 2023, 41% of vascular plant names on the Global Biodiversity Information Facility (GBIF) were digitally undetectable [36]. Open data in databases tend not to be indexed in search engines, and therefore tend to to be undiscoverable digitally [7]. This was apparent when exploring databases: some sources were encountered incidentally through associated websites and papers or identified from the metadata of Brandt et al. [14], rather than retrieved from search results.
Despite recent growth, TRY data still have issues of completeness and representation, especially for continuous (quantitative) traits compared to categorical traits. There is also a regional bias of data towards North America, Europe, East Asia, and Oceania, and against West Asia, Africa, India, and Canada, skewing representation [8], [15]. This, combined with other regional databases found and used covering Australia, the Mediterranean basin, and central Europe, means data presence is likely biased towards plant species native to or present in these locations and against species not present in them [8]. This could result in failure to identify potentially problematic species from locations underrepresented in the data.
The value of open-access databases is well known. Their data can be reused for other analyses, allow for hypothesis testing, improve the cost-effectiveness of measurements, and share efforts among many people [1], [7]. Plant trait databases specifically allow the study of functional features of communities [4], the understanding of global environmental change, and applications to conservation and invasion biology [9]. However, several concerns surround fully open data. Ethical issues with sensitive data, including that which belongs to or impacts Indigenous Peoples, must optimise the CARE principles of Collective benefit, Authority to control, Responsibility, and Ethics, and hence may be unable to be fully open [37]. Additionally, proprietary limitations and resistance from publishers who would benefit from paid access exist [1], [38]. For authors contributing to databases, there is a lack of incentives, infrastructure, and support. Data sharing is undervalued compared to article publication [1], and worries exist concerning misuse, intellectual property, and misinterpretation. Furthermore, data sharing does not benefit contributors as much as consumers [7]. Nevertheless, open access to botanical and ecological data has increased, and continues to do so [15], [38].
Data must be meaningfully accessible to be used by other authors: interpretable with descriptive metadata, and, especially within a database, quality-controlled and standardised [1], [6]. Accessible data must follow the FAIR principles of Findability, Accessibility, Interoperability, and Reusability. These criteria are not always met; historically, data have not been collected with these principles in mind, and the lack of normalisation and standardisation within some life sciences databases has increased the difficulty [7]. This prevented the inclusion of some data; for example, the dispersal mechanism data in the Pannonian Database of Plant Traits lacked descriptive metadata and definitions needed for understanding. On the other hand, TRY data includes original sources, so codified values could be understood through their original source metadata. For plant traits, additional values such as habitat type and methods are also needed for meaningful accessibility [9]. Taxonomy is another issue for meaningful accessibility. Some species are listed only at the genus level, hybrids can be difficult to deal with, misidentification is challenging to fix, and taxonomy is dynamic, so synonyms and redundant names pose issues [11]. As mentioned in the methods section, some issues were encountered with changing taxonomy, and standards varied between different sources. As Kattge et al. note [9], different sources may also use different species concepts.
A further issue with database accuracy and application is that values are often coalesced for each species across different populations, habitats, and sometimes sources [4]. Different sources mean combining inaccurate and accurate data, highlighting the need for quality control [5]. Fortunately, some sources, including TRY, contain multiple values per species per trait, capturing some intraspecific variation [9]. Trait values can change with environmental variability, especially for traits known to be more phenotypically plastic [15]. These traits tend to be quantitative rather than categorical [4]. For example, fruit type was fixed across all sources examined, whereas the main dispersal mode could vary between sources (as also noted by Wotton and McAlpine [39]). Additionally, shade tolerance measurements varied across sources and often between different measurements from the same source. Intraspecific, environment-related variation can increase competitive and invasive ability in a new location or changing climate [40], [41]. For example, the sycamore maple (Acer pseudoplatanus) is relatively shade-intolerant in its native French range, but establishes and survives under closed-canopy forest in Aotearoa (introduced/invasive range), potentially due to a combination of natural release from herbivory, genotypic differences, phenotypic plasticity, and abiotic differences [42]. Therefore, findings from database data should be used in combination with context-specific factors; in this context, the data used for interpretation may need to be filtered by those collected in similar habitats, which would itself reduce the amount of data available.
In conclusion, global and local database usage allowed trait analysis of recently naturalised plant species in Aotearoa, which would otherwise be unfeasible to do in a timely manner. However, this was limited by discoverability, completeness, biases, and meaningful accessibility of databases. The resulting gaps in data found, combined with geographic biases, reduced representativeness, may decrease accuracy and usefulness for the application of data to biosecurity. This highlights the importance of, and the need to continue improving, open data sharing, along with the required infrastructure and incentives to do so.
Acknowledgements
Thank you to my supervisor, Margaret Stanley, for the opportunities, guidance, in-depth and speedy feedback, and great (and hilarious) chats, and to Keiko Hashiba for allowing me to be involved in her PhD research through this project, for the handy sources, direction, and feedback. I wish her well with her ongoing research! I was lucky to be part of the Cain-Stainly lab group this summer; thanks to everyone there, especially for the presentation feedback and fieldwork adventures. Thank you to Bioprotection Aotearoa for the opportunity to take part in your Summer Scholarship Programme.

Figure 2: Sycamore maple (Acer pseudoplatanus) growing in Wellington [43].
Methods
I selected the 70 non-native species that were casual-occurring as of 2006, and were fully naturalised as of 2020, using data from Brandt et al. [14]. For each species, accessible and useful information on dispersal mode, fruit type, and shade tolerance traits was extracted from databases.
TRY Plant Trait Database
Trait ID numbers and names requested from the TRY plant trait database [15] were 28: dispersal syndrome; 99: fruit type; 231: dispersal unit type; 232: dispersal unit shape; 350: seed morphology type; 586: fruit/seed conspicuousness; 603: species tolerance to shade; 788: species light requirement; 813: plant morphological adaptations (seed or dispersal unit metamorphoses); 892: dispersal unit appendages; 1131: species environmental indicator value according to Ellenberg, (light); 1263: dispersal unit floating capacity; 2939: fruit pericarp type; 2940: fruit dehiscence type; 2941: fruit surface type; 3048: fruit palatability; 3600: fruit shape; 3720: species light preference; and 3981: fruit texture inside. TRY species ID numbers for the 70 species were identified, and where unidentifiable, homotypic synonyms were used [16], [17]. An ID number was found for 67 of the 70 species or their synonyms. Both public and private data were requested.
AusTraits Database
From the AusTraits database [18], data associated with the following traits for each species were extracted using R Studio (v4.3.2) [19]: fruit_type, fruit_dehiscence, fruit_fleshiness, fruit_colour, fruiting_time, dispersal_syndrome, dispersal_appendage, dispersal_unit, reproductive_light_environment_index, establishment_light_environment_index, and seedling_establishment_conditions. The APCalign R package (v1.1.3) [20] was used to align species names to current standards. Where a species name was not aligned, homotypic synonyms were tried. Aligned names were found for 68 of the 70 species or their synonyms. The aligned names were then searched for in the AusTraits database using the AusTraits R package (v3.1.1) [18], and 41 of the 70 species were found.
Other Databases and Sources
All data from the BROT 2.0 plant traits database for Mediterranean basin species [21] were downloaded and filtered by taxon and trait names. Of the 44 traits available, the numbers and names of those used were 23: dispersal mode and 26: fruit type.
The “Ellenberg-type indicator values” dataset from the Pladias Database was downloaded [22], [23]. For relevant species present, the L (light requirements) values were filtered. L values (species-specific numeric values) were used instead of Lx values (generalist values) when the two differed. The “Dispersal strategy” dataset was also downloaded and bracketed standard dispersal definitions were used, rather than the other taxon-type categorisation available (e.g., autochory [self-dispersal] rather than ‘allium-type’).
The “EIV L” (Ellenberg’s light indicator) and “Dispersal strategy” datasets were downloaded from the Pannonian Database of Plant Traits [24], [25]. Dispersal mechanisms were discarded because they were not in an interpretable format. EIV L values were accessible.
All species with missing shade-tolerance data were searched for in the Plants For a Future database [26]. The database search page was used instead of purchasing access to the database spreadsheet, as the shade-tolerance data available were assumed to be the same based on sample database pages.
The following web pages were searched for some species with remaining gaps: Australian Tropical Rainforest Plants, Conservation Auckland, North Carolina Extension Gardener Plant Toolbox, New Zealand Plant Conservation Network, PlantZAfrica, Weeds Australia, and World Flora Online [27] - [33]. Two family-wide generalisations for fruit type were made based on Veldkamp et al. [34] and Reutemann et al. [35].
For each trait, information was transcribed if it explicitly gave a value or provided information that could be used to determine the trait value. To form a condensed dataset, trait-specific values were set, and transcribed information was used to determine appropriate values for each species with available data.
Results
Trait data were found for between 50.0% and 78.6% of the 70 species, depending on the trait. Shade tolerance was available for the lowest proportion of species, and fruit type for the highest. None of the three trait values were found for 14.3% of species, and all three trait values were found for 37.1% of species.
[1] D. Kapiszewski and S. Karcher, “Making research data accessible,” in The Production of Knowledge, 1st ed., C. Elman, J. Gerring, and J. Mahoney, Eds., Cambridge, UK: Cambridge University Press, 2020, pp. 197–220, doi: 10.1017/9781108762519.008.
[2] S. S. Gantayat, A. Misra, and B. S. Panda, “A Study of Incomplete Data – A Review,” in Proceedings of the International Conference on Frontiers of Intelligent Computing: Theory and Applications (FICTA), 2013, pp. 401–408, doi: 10.1007/978-3-319-02931-3_45.
[3] L. Antoniuk and D. Taskaiev, “Concept, types and importance of databases. Prospects for their development,” in Proceedings of the 8th International Scientific and Practical Conference, Mar. 2023, pp. 474–480. [Online]. Available: https://archive.interconf.center/index.php/conference-proceeding/article/view/2866.
[4] V. Cordlandwehr, R. L. Meredith, W. A. Ozinga, R. M. Bekker, J. M. Van Groenendael, and J. P. Bakker, “Do plant traits retrieved from a database accurately predict on‐site measurements?,” J. Ecol., vol. 101, no. 3, pp. 662–670, May 2013, doi: 10.1111/1365-2745.12091.
[5] Á. Enríquez-de-Salamanca, “Botanical databases in EIA: Opportunities and challenges,” Impact Assess. Proj. Apprais., vol. 43, no. 4, pp. 302–312, Jul. 2025, doi: 10.1080/14615517.2025.2482137.
[6] D. P. C. Peters, “Accessible ecology: synthesis of the long, deep, and broad,” Trends Ecol. Evol., vol. 25, no. 10, pp. 592–601, Oct. 2010, doi: 10.1016/j.tree.2010.07.005.
[7] A. E. Thessen and D. L. Patterson, “Data issues in the life sciences,” ZooKeys, vol. 150, pp. 15–51, Nov. 2011, doi: 10.3897/zookeys.150.1766.
[8] Y. Lin, Y. Guan, A. Asudeh, and H. V. Jagadish, “Identifying insufficient data coverage in databases with multiple relations,” Proc. VLDB Endow., vol. 13, no. 12, pp. 2229–2242, Aug. 2020, doi: 10.14778/3407790.3407821.
[9] J. Kattge et al., “A generic structure for plant trait databases,” Methods Ecol. Evol., vol. 2, no. 2, pp. 202–213, Apr. 2011, doi: 10.1111/j.2041-210X.2010.00067.x.
[10] J. A. Williams and C. J. West, “Environmental weeds in Australia and New Zealand: issues and approaches to management,” Austral Ecol., vol. 25, no. 5, pp. 425–444, Oct. 2000, doi: 10.1046/j.1442-9993.2000.01081.x.
[11] K. G. McAlpine and C. J. Howell, “List of environmental weeds in New Zealand 2024,” Department of Conservation, Wellington, New Zealand, Science for Conservation 340, 2024. [Online]. Available: https://www.doc.govt.nz/globalassets/documents/science-and-technical/sfc340.pdf.
[12] M. Stanley and I. Bassett, “Environmental weeds in New Zealand: Impacts and management,” in Austral Ark: The State of Wildlife in Australia and New Zealand, 1st ed., A. Stow, N. Maclean, and G. I. Holwell, Eds., Cambridge, UK: Cambridge University Press, 2014, pp. 134–161, doi: 10.1017/CBO9781139519960.
[13] A. K. Cummack and M. Hassan, Global Databases. 2026. [Digital Image].
[14] A. Brandt, H. Maule, and J. Fridley, “NZ non-native flora growth form data 2020.” DataStore: The Manaaki Whenua - Landcare Research Data Repository, 2020. [Online]. Available: https://datastore.landcareresearch.co.nz/en_AU/dataset/nz-non-native-flora-growth-forms-2020/resource/80532772-65f0-449e-850f-c3607063c312
[15] J. Kattge et al., “TRY plant trait database – enhanced coverage and open access,” Glob. Change Biol., vol. 26, no. 1, pp. 119–188, Jan. 2020, doi: 10.1111/gcb.14904.
[16] R. Govaerts, E. Nic Lughadha, N. Black, R. Turner, and A. Paton, “The world checklist of vascular plants, a continuously updated resource for exploring global plant diversity,” Sci. Data, vol. 8, no. 1, p. 215, Aug. 2021, doi: 10.1038/s41597-021-00997-6.
[17] Royal Botanic Gardens, Kew, “Plants of the World Online.” Accessed: Feb. 11, 2026. [Online]. Available: https://powo.science.kew.org/
[18] D. Falster et al., “AusTraits, a curated plant trait database for the Australian flora,” Sci. Data, vol. 8, no. 1, p. 254, Sep. 2021, doi: 10.1038/s41597-021-01006-6.
[19] R Core Team, “The R Project for Statistical Computing.” R Foundation for Statistical Computing. [Online]. Available: https://www.R-project.org/.
[20] E. Wenk et al., “APCalign: an R package workflow and app for aligning and updating flora names to the Australian Plant Census,” Aust. J. Bot., vol. 72, no. 4, Jun. 2024, doi: 10.1071/BT24014.
[21] J. G. Pausas and Ç. Tavşanoğlu, July 2018, “BROT plant functional trait database: Data file,” doi: 10.6084/M9.FIGSHARE.5280868.
[22] M. Chytrý, L. Tichý, P. Dřevojan, J. Sádlo, and D. Zelený, “Ellenberg-type indicator values for the Czech flora,” Preslia, vol. 90, no. 2, pp. 83–103, May 2018, doi: 10.23855/preslia.2018.083.
[23] M. Chytrý et al., “Pladias Database of the Czech flora and vegetation,” Preslia, vol. 93, no. 1, pp. 1–87, 2021, doi: 10.23855/preslia.2021.001.
[24] Pannonian Database of Plant Traits, “Dispersal strategy.” padapt.eu. Accessed: Feb. 11, 2026. [Online]. Available: https://padapt.eu/download.html.
[25] Pannonian Database of Plant Traits, “EIV L.” padapt.eu. Accessed: Feb. 11, 2026. [Online]. Available: https://padapt.eu/download.html.
[26] Plants For A Future, “Database,” pfaf.org. Accessed: Feb. 11, 2026. [Online]. Available: https://pfaf.org/user/DatabaseSearhResult.aspx.
[27] Centre for Australian National Biodiversity Research, Australian Tropical Herbarium, and Australian Biological Resources Study, “Browse species,” Australian Tropical Rainforest Plants. Accessed: Feb. 01, 2026. [Online]. Available: https://apps.lucidcentral.org/rainforest/text/entities/index.htm.
[28] Tiaki Tāmaki Makaurau Conservation Auckland, “Pest Search,” Auckland Council. Accessed: Feb. 11, 2026. [Online]. Available: https://www.tiakitamakimakaurau.nz/protect-and-restore-our-environment/pests-in-auckland/pest-search.
[29] NC Cooperative Extension and NC State University, “Find a Plant,” North Carolina Extension Gardener Plant Toolbox. Accessed: Feb. 11, 2026. [Online]. Available: https://plants.ces.ncsu.edu/find_a_plant/.
[30] New Zealand Plant Conservation Network, “New Zealand’s Flora,” nzpcn.org.nz. Accessed: Feb. 11, 2026. [Online]. Available: https://www.nzpcn.org.nz/flora/.
[31] South African National Biodiversity Institute, “PlantZAfrica.” Accessed: Feb. 11, 2026. [Online]. Available: https://pza.sanbi.org/.
[32] Centre for Invasive Species Solutions, “Weeds Australia.” Accessed: Feb. 11, 2026. [Online]. Available: https://weeds.org.au/.
[33] World Flora Online, “Classification,” WFO: The World Flora Online. Accessed: Feb. 11, 2026. [Online]. Available: https://www.worldfloraonline.org/classification.
[34] J. F. Veldkamp, H. Duistermaat, K. M. Wong, and D. J. Middleton, “Poaceae,” in Flora of Singapore, vol. 7, Singapore Botanic Gardens, 2019, pp. 219–501. doi: 10.26492/fos7.2019-07.
[35] A. G. Reutemann, J. A. B. San Martin, and R. E. Pozner, “Structural and histochemical approach to the fruit and seed diversity of Cyperaceae in an evolutionary context,” Plant Reprod., vol. 37, no. 2, pp. 147–170, Jun. 2024, doi: 10.1007/s00497-023-00465-8.
[36] N. Nicolson, M. Trekels, Q. J. Groom, S. Knapp, and A. J. Paton, “Global access to nomenclatural botanical resources: Evaluating open access availability,” Plants People Planet, vol. 5, no. 6, pp. 899–907, Nov. 2023, doi: 10.1002/ppp3.10438.
[37] S. R. Carroll, E. Herczog, M. Hudson, K. Russell, and S. Stall, “Operationalizing the CARE and FAIR Principles for Indigenous data futures,” Sci. Data, vol. 8, no. 1, p. 108, Apr. 2021, doi: 10.1038/s41597-021-00892-0.
[38] S. Matutinovic, “Open access in botany,” Bot. Serbica, vol. 38, no. 1, pp. 191–195, 2014, [Online]. Available: https://botanicaserbica.bio.bg.ac.rs/arhiva/pdf/2014_38_1_610_full.pdf.
[39] D. M. Wotton and K. G. McAlpine, “Seed dispersal of fleshy-fruited environmental weeds in New Zealand,” N. Z. J. Ecol., vol. 39, no. 2, pp. 155–169, 2015. [Online]. Available: https://www.jstor.org/stable/10.2307/26198707.
[40] C. S. Sheppard, B. R. Burns, and M. C. Stanley, “Future-proofing weed management for the effects of climate change: Is New Zealand underestimating the risk of increased plant invasions?,” N. Z. J. Ecol., vol. 40, no. 3, pp. 398–405, 2016, doi: 10.20417/nzjecol.40.45.
[41] D. M. Wotton and K. G. McAlpine, “Predicting native plant succession through woody weeds in New Zealand,” Department of Conservation, Wellington, New Zealand, Research and Development Series 336, 2013. [Online]. Available: https://www.doc.govt.nz/globalassets/documents/science-and-technical/drds336entire.pdf.
[42] S. Shouman et al., “Functional shift of sycamore maple (Acer pseudoplatanus) towards greater plasticity and shade tolerance in its invasive range,” Perspect. Plant Ecol. Evol. Syst., vol. 29, pp. 30–40, Dec. 2017, doi: 10.1016/j.ppees.2017.11.001.
[43] J. R. Rolfe, Wellington. 2008. Accessed: Mar. 13, 2026. [Photo]. Available: https://www.nzpcn.org.nz/flora/species/acer-pseudoplatanus/

Amie is in their final year of undergraduate study, and they can’t stay away from research projects. She is a plant nerd, passionate about environmental and human rights, and has research interests at the intersections between sciences, the social sciences, the humanities, and politics.